

Our interests focus on the functions of Girdin (reviewed in Wang et al., Pathol Int, 2016) and CD109 (reviewed in Mii et al., Pathol Int, 2019) that we previously identified as the downstream effectors of the Ret signaling pathway.
Girdin, which is also known as Gα-interacting vesicle-associated protein (GIV), is an actin-binding and hub protein with many interacting proteins, and is involved in cell migration, polarization, cargo-selective endocytosis, and metabolism (Fig 1; Dev Cell, 2005; Cancer Res, 2008; Nat Cell Biol, 2008, Neuron, 2009; Oncogene, 2012; EMBO J, 2014; Cancer Res, 2015; PLoS Biol, 2018). In addition, the results of our studies on Girdin-deficient and mutant mice and human tissue samples have suggested that it is an essential regulator of collective migration and invasion of neuroblasts in the brain and cancer cells, respectively (J Neurosci, 2011; Cancer Sic, 2018, and others). We also identified that DISC1, which is encoded by a schizophrenia susceptibility gene, interacts with Girdin, and the DISC1-Girdin complex regulates the migration and positioning of newly generated neuroblasts in the hippocampus (Neuron, 2009; J Neurosci, 2011). The analysis of Daple, which is a member of the Girdin family of proteins, has also been the scope of research in our laboratory (Nat Commun, 2012; Cell Rep, 2014).
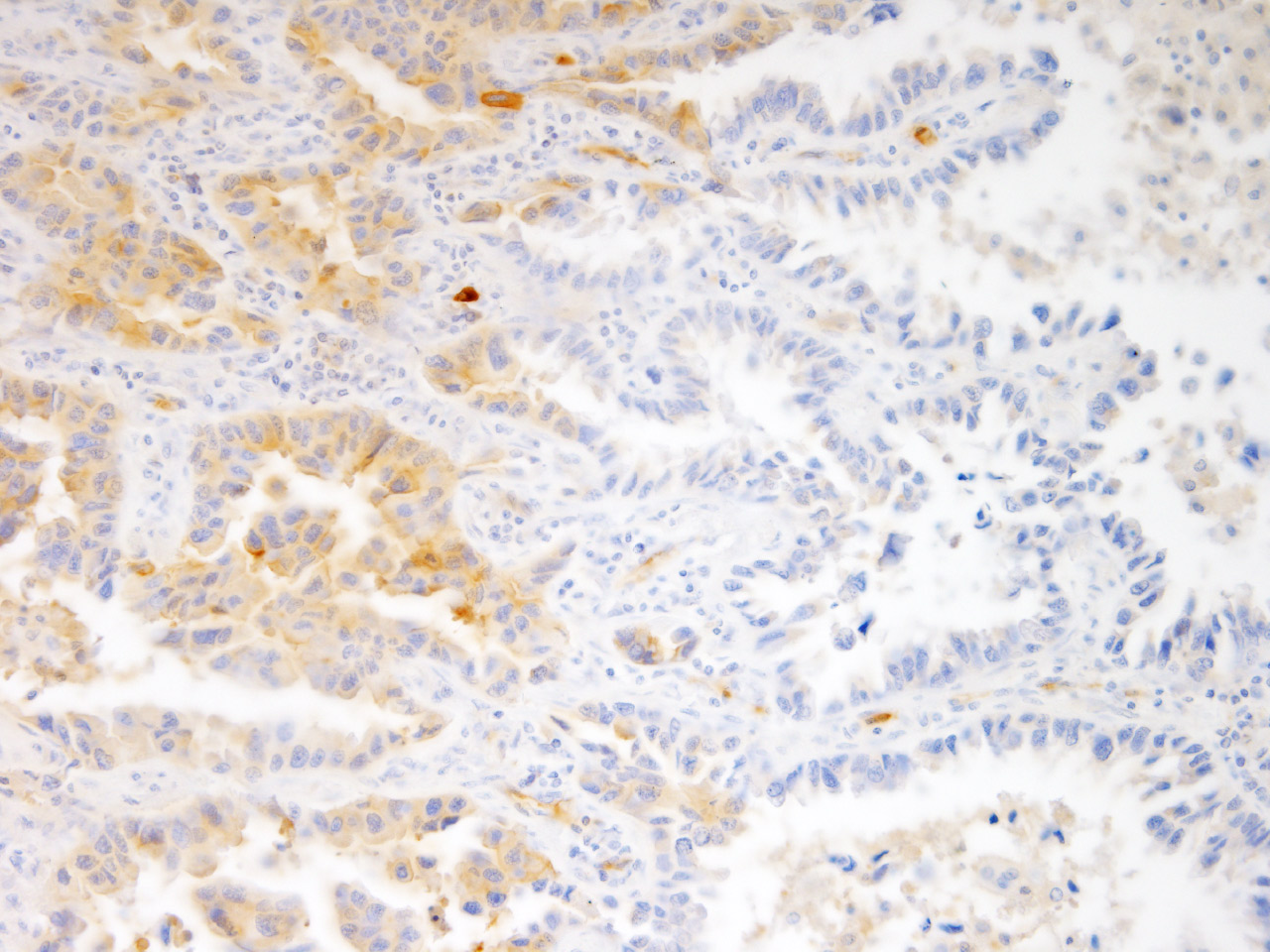
CD109 is a GPI-anchored membrane protein, which is expressed in the basal cells of the skin as well as in the myoepithelial and endothelial cells in normal tissues. It is highly overexpressed in many types of cancer cells, in which it binds to the TGF-β receptors and matrix proteins to regulate intracellular signals (Pathol Int, 2008; Oncogene, 2010; Am J Pathol, 2012; PLoS One, 2014; Oncotarget, 2016). CD109 is also highly expressed in the brain tumor stem cells (BTSCs) of gliomas and is essential for the maintenance of stemness of BTSCs (J Pathol, 2017). Our recent observation indicated that CD109 is a marker of invasive cancer cells in human lung adenocarcinoma, wherein it regulates the activation of the latent form of TGF-β in the tumor stroma (Fig 2). Owing to these findings, CD109 is one of the molecular targets for developing new anticancer therapeutics and has attracted increasing attention in the pharmaceutical sectors.
We will continue to delve into the molecular functions of these proteins and their family of proteins and reveal their involvement in the physiology of multiple tissues and the pathology of human diseases.
Fig. 1
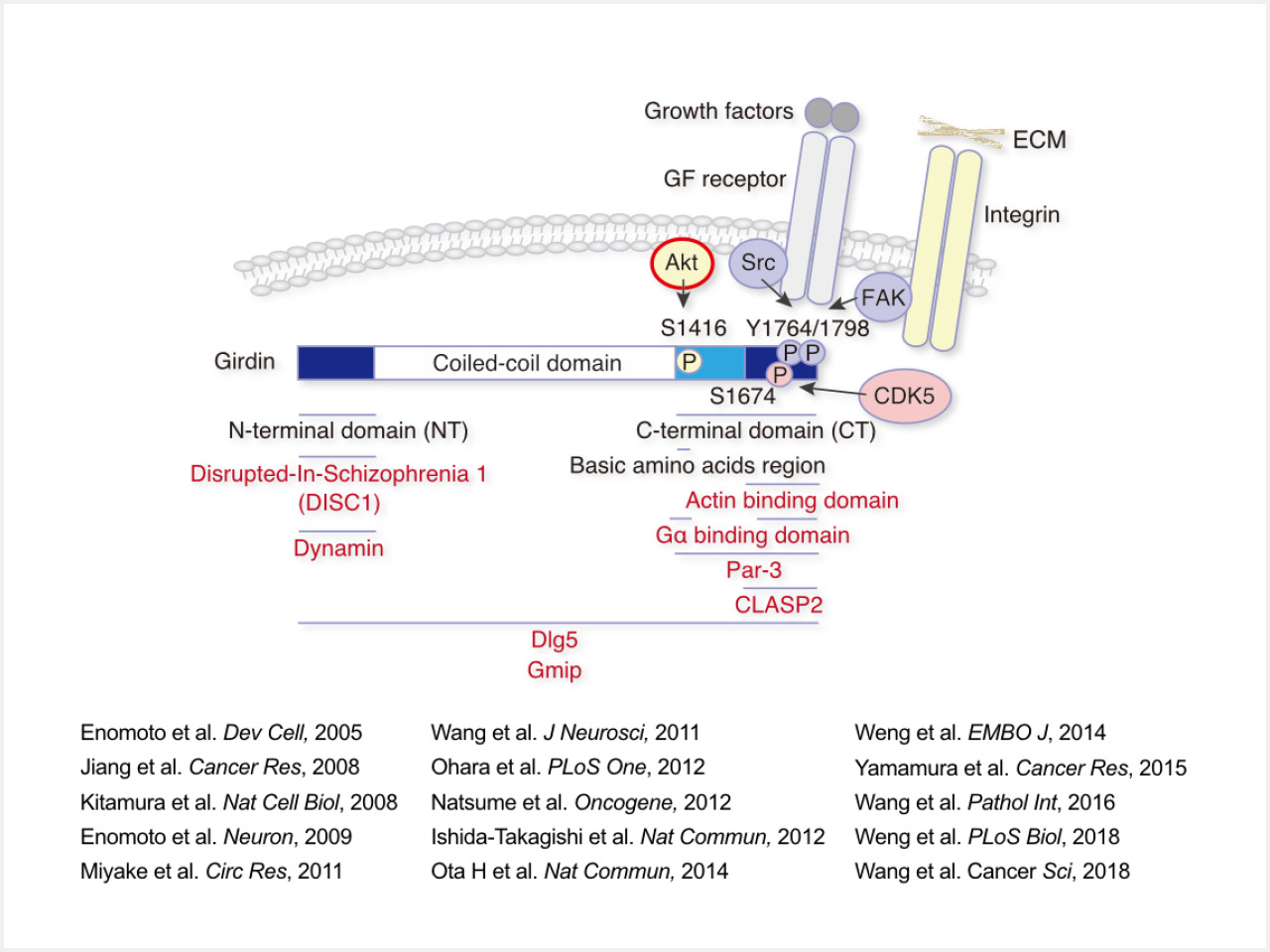
Fig. 2
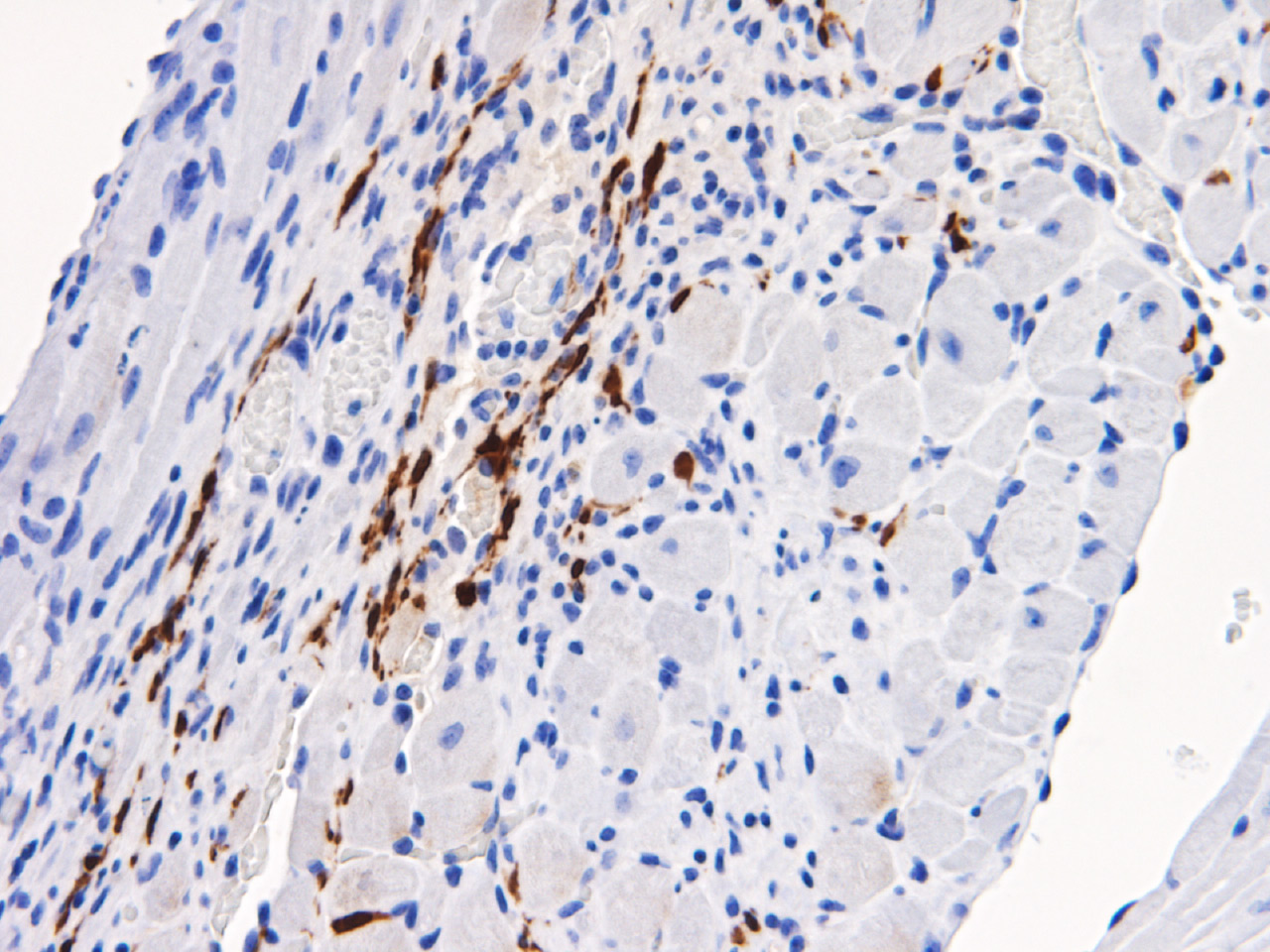
Cancer stroma comprises multiple components, including cancer-associated fibroblasts (CAFs), tumor vessels, immune cells, and the extracellular matrix. CAF proliferation is particularly prominent in devastating cancers such as pancreatic cancer. Fibroblast proliferation and alterations in its phenotype are also major causes of decline in organ functions that are associated with aging, chronic inflammation, and tissue wound and repair. Recent evidence suggests that in the case of cancer and fibrotic diseases, the stroma contains heterogeneous populations of fibroblasts (reviewed in Kobayashi et al., Nat Rev Gastroenterol Hepatol, 2019). We recently identified a GPI-anchored membrane protein, Meflin, that is a marker protein of cancer-restraining CAFs (rCAFs) and fibroblasts that inhibit fibrosis. Meflin plays a suppressive role in the progression of pancreatic cancer (Fig 3; Cancer Res, 2019; reviewed in Miyai et al., Cancer Sci, 2020) and is essential for tissue repair after acute myocardial infarction (Fig 4; Circ Res, 2019). Interestingly, induction of heart failure in Meflin-deficient mice resulted in the development of heart failure with preserved ejection fraction (HFpEF), the prevalence of which has shown an increase in the clinic recently. HFpEF is characterized by the stiffening of the heart tissue and diastolic dysfunction (Circ Res, 2019). Our next goal is to continue analyzing the significance of fibroblast heterogeneity in various human diseases by focusing on Meflin and other fibroblast markers.
Fig. 3
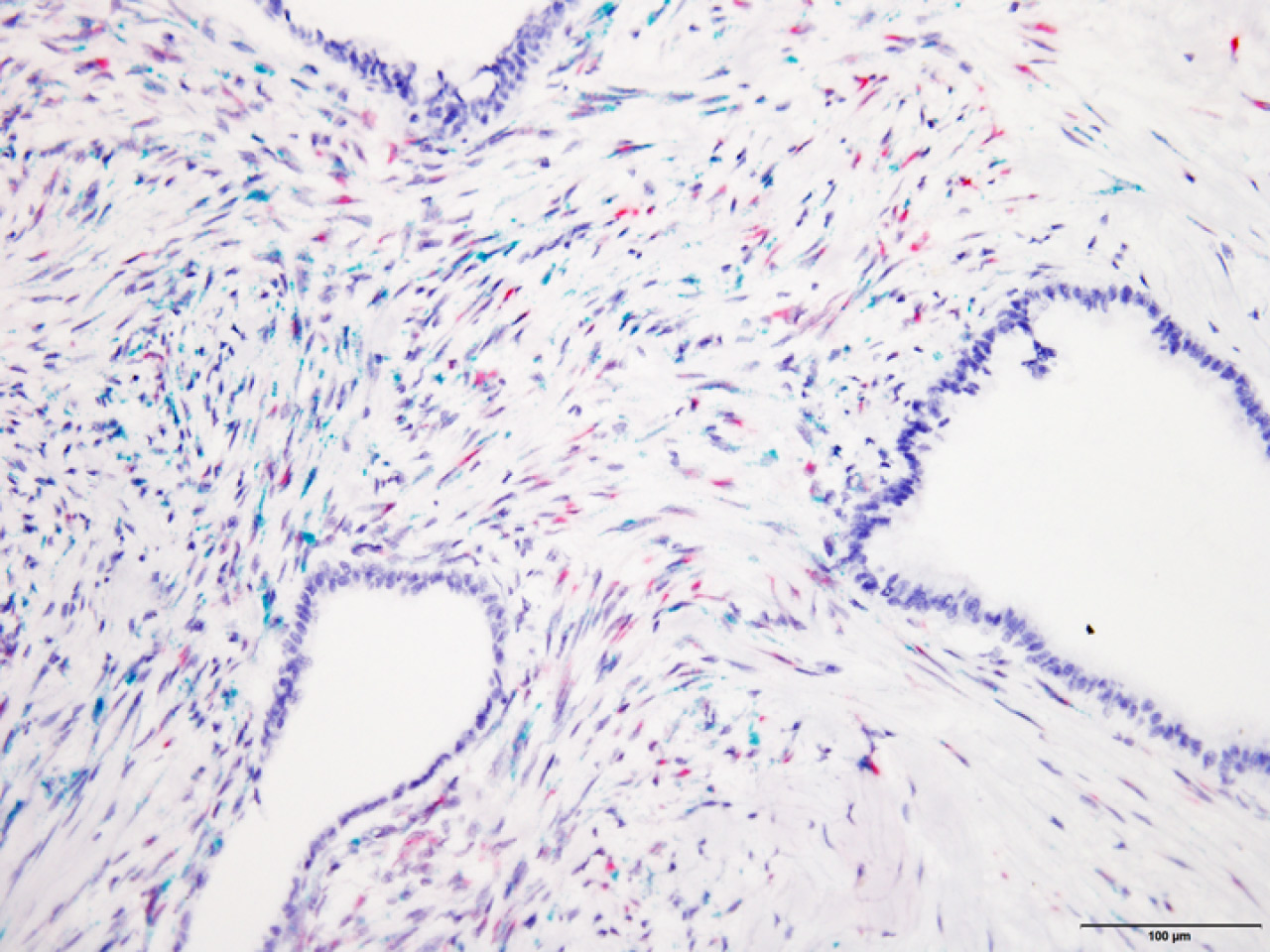
Fig. 4
Meflin was originally identified as a marker of mesenchymal stem/stromal cells (MSCs) (Sci Rep, 2016). Notably, its expression is limited to undifferentiated MSCs, but is absent in differentiated MSC lineages such as osteocytes, chondrocytes, adipocytes and muscle cells, and this clearly differentiates it from other conventional MSC markers such as CD90, CD105, CD73, and CD44. As described above, Meflin is also expressed in a fibroblast subset that is located in the perivascular areas in multiple tissue types. Although fibroblasts are well known as an essential architectural component that maintains the structure of every organ and tissue, results of various recent studies revealed that some of them function as niches for tissue stem cells and are actively involved in the physiological functions of multiple organs. However, in disease conditions, the changes in the fibroblast phenotype are accompanied by the production of the extracellular matrix, which results in the stiffening of the tissues. Importantly, the mechanical stress induced by this stiff environment further alters fibroblast function along with its gene expression. Results of our recent investigation on the gene expression profile of fibroblasts indicated that the nature of fibroblasts is not completely understood yet and is much more complicated than expected. Therefore, in future, our aim is to address the question “what are fibroblasts?” based on the analysis of the markers of MSCs and fibroblasts. Another scope of our lab will be understanding the mechanisms of human diseases from a viewpoint that the mechanical stress experienced by cells in the stiff tissues is crucial for changes in cell functions, which is known as mechanopathology.